標題 : Spatiotemporal analysis of human intestinal development at single-cell resolution
期刊:Cell
影響因子:41.58
文章導讀
人類對腸道發育的認知還不夠完善,如果將單細胞測序和空間轉錄組學聯合起來,人們就可以兼顧單細胞的分辨率與空間位置的準確性,從而描述腸道發育隨時間變化的形態。本實驗共確定了 101 種細胞形態,包括上皮和間充質細胞群和關鍵形態改變之間的相關性。實驗人員描述了隱窩絨毛軸是如何形成的,以及神經、血管、間充質形態發生和發育中的腸道免疫群體。同時,研究人員還鑒定了發育中的成纖維細胞和肌成纖維細胞亞型的分化層次,并描述了這些亞型的不同功能。此外,他們還發現了派伊爾結和腸道相關淋巴組織(GALT)的起源,并描述了位置特異性免疫程序。利用實驗得出的結論,他們提出了一個形態形成梯度的無偏性分析,直接關聯連續的幾波細胞分化,并確定了細胞及位置與罕見的發育性腸道疾病有關。
科學問題
腸道有著怎樣的發育過程?
腸發育過程的紊亂又怎樣與疾病相關?
實驗設計
從 17 個胚胎中不同發育時間點與組織位置取出的共 77 個腸道樣本,總計 76592 個細胞。
技術:10X genomics
實驗思路
實驗結果
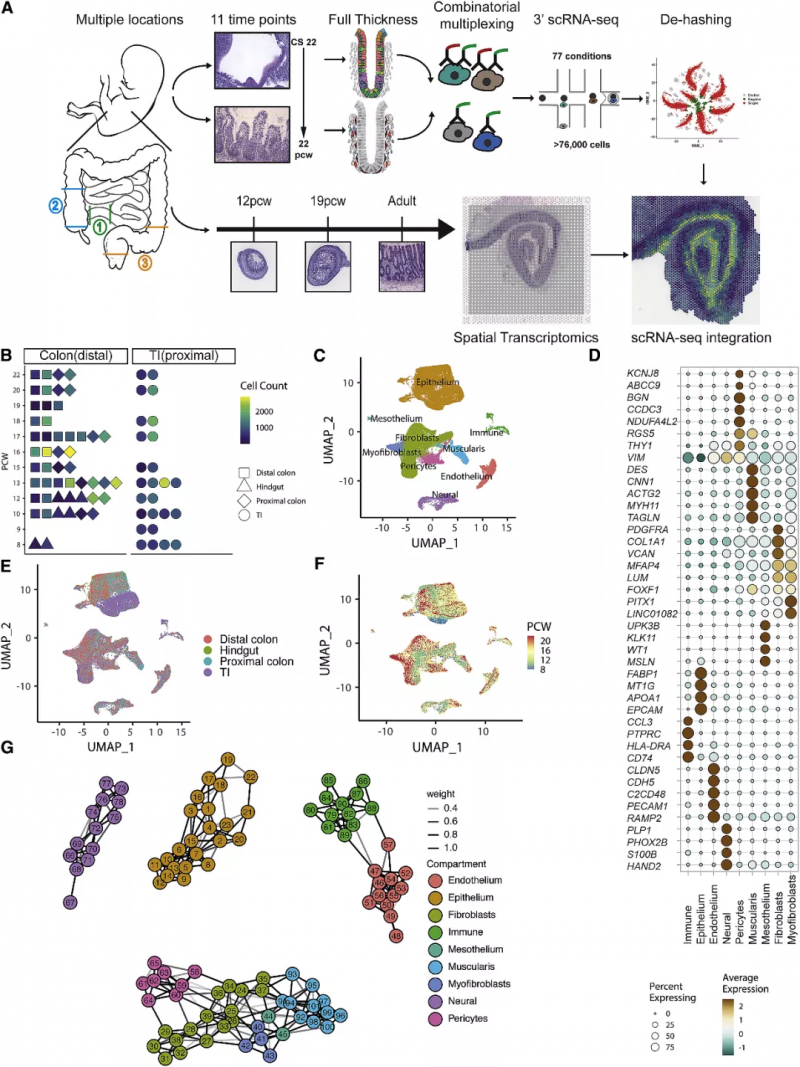
圖 1 人類腸道發育的時空轉錄圖譜
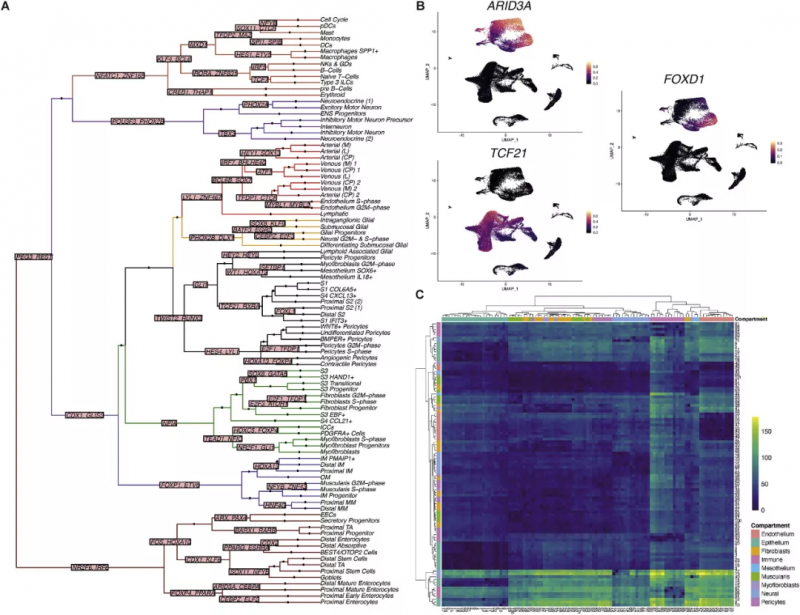
圖 2 胎兒腸道發育的轉錄因子調控網絡和受體 - 配體相互作用
1、根據發育時間和空間對 101 種腸細胞的分類
作者從代表不同發育時間點和組織位置的 17 個胚胎收集的 77 個腸道樣本中 10x Genomics 的單細胞基因表達解決方案生成了 scRNA 序列圖譜(圖 1),經過聚類,分為 9 種腸道組成細胞(圖 1C,D),并展示了它們在空間位置與發育時間的差異(圖 1E,F)。通過進一步的細分,基于標志基因劃分出共 101 個細胞亞群(圖 1G)。接下來研究人員又基于 464 種目前已知的發育轉錄因子針對細胞命運繪制了“決定樹”(圖 2A),并對 101 個亞群的細胞進行了細胞通訊分析(圖 2C),辨認假定的受體 - 配體對。
2、單細胞的空間位置
為了繪制 scRNA-seq 數據的空間分布圖,作者利用 10x Genomics 的 Visium 空間轉錄組技術對來自整個腸道發育的組織進行了空間轉錄組學分析,在每張切片中確定了映射到離散位置的 5 -13 個點簇,以上述 scRNA-seq 圖譜為參考,作者通過因子分析以確定每個聚點可能的單細胞組成,從而在空間上定位所有 scRNA-seq 簇(圖 3A)。這種定位能夠很好地描述細胞類型,例如在隱窩底部的上皮干細胞(ISCs)(圖 3Ai 與 3Aii),位于平滑肌外層的 FOXF2- 肌層細胞(圖 3Aiii),以及處于隱窩頂部的 BEST4/OTOP2 細胞(圖 3Aiv)。圖 3B 展示了隱窩頂部腸細胞與肌成纖維細胞合適的解剖學定位,RET 在肌腸叢特異表達,PTPRC (CD45) 在粘膜下淋巴濾泡特異表達。
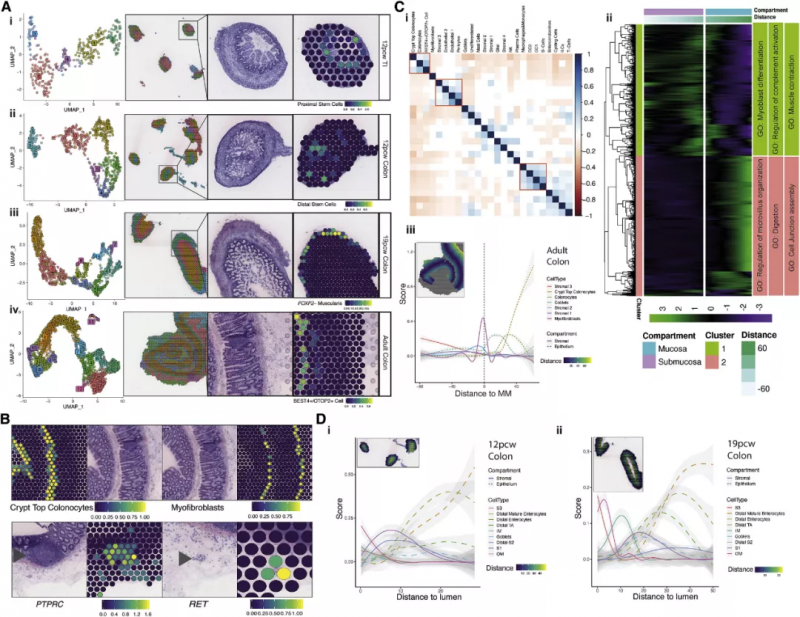
圖 3 ST 和 scRNA-seq 整合的腸道發育時空分析
3、人類腸上皮細胞與 ISCs 的發育
根據細胞的吸收性、分泌性、未分化與干細胞基因特征可以很輕易地對 17622 個被捕獲的上皮細胞進行區分(圖 4A)。分析結果表明近端(小腸 [SI]) 和遠端(結腸)樣本之間有著顯著位置差異,這一差異由高特異性基因表達(如 CCL25 和 APOE) 劃分(圖 4A 和 4B)。這表明,在隱窩形成之前,特定位置的轉錄程序已經在發育中建立起來。隨著時間的推移,可以觀察到上皮細胞的大量細胞重塑,在 12 PCW(post-conceptual weeks)之前,祖細胞 /TA 細胞占主要組成部分,很少看到成熟的吸收和分泌細胞,而在 12PCW 之后,上皮組織的組成已經與成人組織相似(圖 4C)。在對 ISCs 發育的研究過程中,實驗人員發現了一群近端上皮干細胞樣祖細胞,進而會形成早期腸細胞。這些細胞表現出許多原始特征,包括在中胚層分化中重要的 VTN 高表達,與 ISCs 相比,LGR5 表達很少,ONECUT2 表達參與上皮發育(圖 4B)。在小腸中,這些細胞獨特地表達轉錄因子 GATA4(圖 4E),在 12 周之后,這一表達又大部分丟失。這些細胞高度表達轉鐵蛋白(圖 4B,F),證實了在絨毛形成過程中鐵代謝的重要性。LGR5 是 ISC 的特征基因,在妊娠早期(<12PCW)被檢測出在近端 / 遠端 ISCs 及干細胞樣祖細胞中廣泛的低表達,即使在隱窩形態建立后(例如,19 個 PCW 后),ISCs 在遠端 / 近端樣本中分別占捕獲的上皮細胞的平均比例為 18%-22%,高于成人結腸 scRNA-seq 研究中捕獲的 3%-4%。由此實驗人員發現了隨發育進行的 ASCL2 轉錄因子模塊的顯著表達上升(包含下游調控目標 LGR5)。
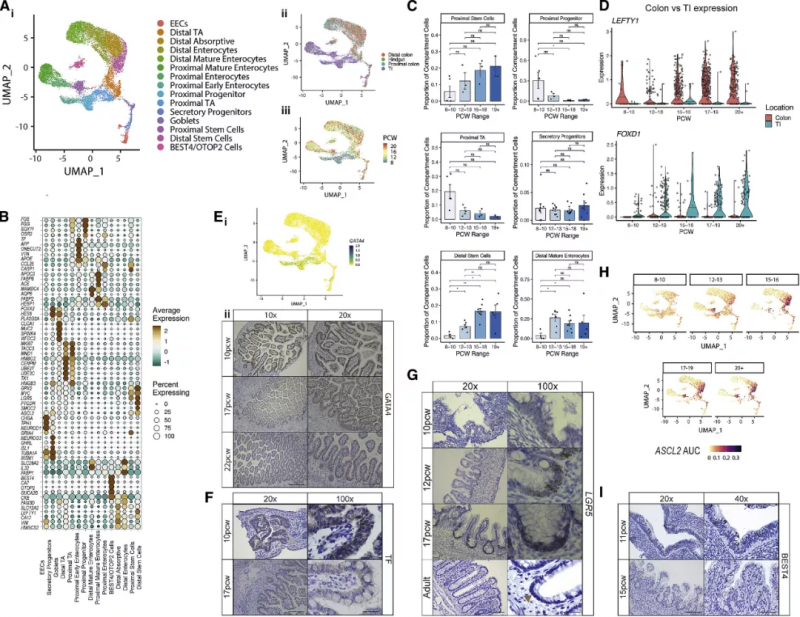
圖 4 子宮上皮成熟和隱窩發育
4、腸道血管生成,神經系統和肌層固有層發生,腸固有層間充質細胞發育
如圖 5A 所示上皮細胞被劃分為靜脈型,動脈型和淋巴型細胞,根據血管的尺寸,它們又被進一步細分。隨著發育時間進程可以觀察到小血管內皮細胞向大血管內皮細胞的轉變,反映了腸道血管發生,驅動這一過程的轉錄因子網絡已得到辨認,如動靜脈分化因子 HEY1 和 SOX13。作為內皮細胞的補充,周細胞亞型由血管生成驅動因子(PRRX1, THBS4, and ANGPT2) 定義(圖 5B)。實驗人員在腸道發育的早期階段發現了不同的神經元和膠質祖細胞,共捕獲到了 5 種膠質細胞和 7 類神經細胞(圖 5E)。在成年人的腸道內,神經叢被肌肉包圍,圖 5F 中祖細胞和分化的腸平滑肌細胞(iSMCs) 分別用 PLPP2 和 ACTA2 基因區分。他們在腸道發育過程中發現了關鍵的轉錄因子調控網絡:KLF7 存在于肌細胞中,TWIST2 存在于間質和外層肌細胞中,FOXF2(該基因缺失突變會致死)特異性活化于內層肌細胞中。這一發現與平滑肌介導的機械力啟動絨毛形成的可能性相一致。受測細胞中大的類群為間充質細胞,它能夠支持上述的其它細胞組分。基于先前的研究結果這些細胞可被劃分成 s1-s4 四個類別,再輔以時間、空間、分化周期信息,這些細胞進一步被劃分為 16 個類群(圖 5G)。圖 5H 展示了對 s3 細胞類群的空間定位,并對幾種 s3 類群的差異基因表達進行了定位,如 C7(圖 5Hii)。將胎兒細胞類型標志基因對應到成人空間轉錄組切片上,實驗人員發現 S3 HAND1+ 和 S3 EBF+ 細胞聚集在大血管周圍(圖 5Hiii-Hiv),概括了成年人的 S3 類群。這突出了這些細胞可能的作用,形成一個支持腸道血管的生態位。細胞通訊分析證實了這一點,ECs、周細胞和“S3”型成纖維細胞信號在同一空間位點相關(圖 3Ci)。
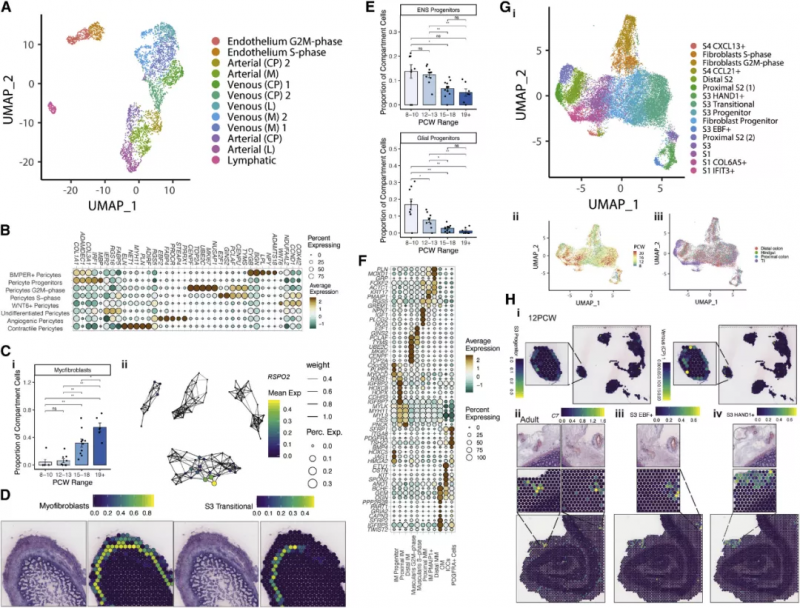
圖 5 間充質和內皮間室細胞的協調發展
5、腸道發育過程中無偏差形成素梯度作圖
圖 6A 中展示了形成素分子的互作關系,虛線標注了富集的信號通路。基因共表達分析辨認出 11 種特異細胞類型和 13 種空間共定位形成素模塊。空間模塊 3 由內皮細胞、成纖維細胞和周細胞來源的形成素分子組成,包括 LRP1,位于組織深部(圖 6Bi)。RSPO3 可以通過 LGR5 向 ISCs 發出信號,它在間皮 / 肌層模塊中較高,在 PCW 12 之前主要可見,然后隨著組織深度的增加而消失(圖 6C)。這說明發育過程存在這樣一種模式:隨著發育中的腸道的生長,形成素分子的梯度可能會因層間物理距離的增加而被打破,但可以通過順序發育的細胞類型的表達來恢復。
6、S2 成纖維細胞具有空間位置特異性并控制上皮模式
實驗人員在發育過程中發現了三個不同的 S2 簇,它們表達 TGF-β (BMP2, BMP4 和 BMP5) 和非標準 WNT 通路(WNT5A 和 WNT5B) 的關鍵形成素分子。這些 s2 特異性基因構成了主要的黏膜下成纖維細胞形成素模塊的一部分,該模塊包含 DLL1、BMP5 和 NRG1(圖 6D)。再加上 RL 相互作用,如 DLL1-NOTCH2,突出了 S2 細胞及其在上皮形成中提供的豐富形態形成生態位的重要性。末端回腸(TI)和結腸 S2 細胞有著 885 個差異表達的基因。在 10 PCW 之前這些細胞展現出許多位點特異性差異,代表了在隱窩 / 絨毛形成之前很強的位置一致性,例如,POSTN 和 BMP3 在它們各自的 S2 亞型中很早就出現了(圖 6E)。S2 形態形成譜中這種關鍵的位置差異指明了一種機制,通過這種機制,可以形成不同的上皮形態。
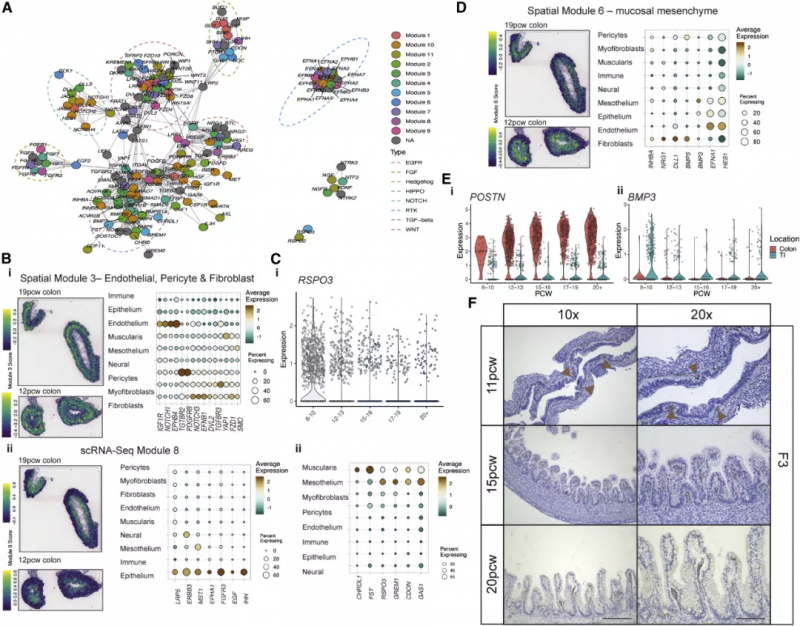
圖 6 在特定的細胞類型和空間位置中的腸道形態發生素梯度
7、人類腸道免疫系統的發育
實驗中一共捕獲到了 6 種共計 2199 個免疫細胞(圖 7A,7B)。在 10 PCW 之前,髓系免疫細胞更為豐富,而在第 12PCW 之后會出現幼稚 CD4+ 和 CD8+ T 細胞、自然殺傷細胞(NK)、1 型先天淋巴細胞(ILCs) 和 3 型 ILCs(圖 7C)。CCL19, CCL21, 與 CXCL13 是和派伊爾結形成有關的基因,并表達于基質組織細胞中。實驗結果表明這些細胞特異性地定位在兩個 S4 成纖維細胞簇中,并且隨著發育表達增加。通過將黏膜下淋巴濾泡作為獨特的結構,研究其空間轉錄組切片,研究人員在這些濾泡內及周圍找到了成人免疫細胞以及 S4 類型細胞(7F),同時也在其中找到了受體 - 配體對,包括 CCR7/CC19。因此認定 S4 細胞是成人結腸中免疫濾泡鄰近的成纖維細胞,在胎兒 GALT 發育過程中以時間依賴性的方式出現。考慮到 S4 CXCL13+ 亞型成纖維細胞的缺失會損害派伊爾結的形成,綜上認定派伊爾結中隱窩周圍成纖維細胞為上皮隱窩生態位支持細胞、淋巴組織形成的協調者和基質 - 免疫交互的介質,實驗結果揭示了它們迄今為止未被認識到的高度動態的作用。
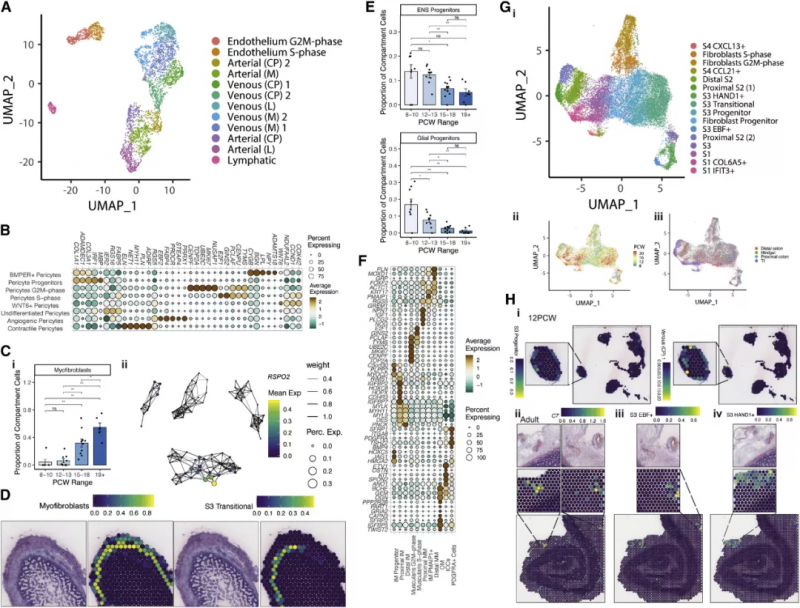
圖 7 早期腸道免疫定植和免疫基質相互作用
8、繪制先天性腸道疾病的細胞基礎圖
為揭露嚴格遵循時間順序的致病轉錄缺陷,實驗人員整合了健康人類與患病者的數據。通過整合 749 個已知的疾病基因和本次實驗的 scRNA-seq 數據,他們將先天性疾病與可能通過高度細胞類型特異性缺陷表現出來并導致腸道、腹側、會陰、神經節、炎癥或腫瘤病理的紊亂的表型聯系起來(圖 8A,B)。SOX10 和 RET 與巨結腸病相關,在腸神經系統中表達(圖 8Cii),包括淋巴相關膠質細胞。考慮到這些細胞在淋巴濾泡形成中發揮作用,實驗數據提示了先天性巨結腸疾病與并發癥小腸結腸炎的潛在聯系——這種并發癥是由神經節腸切除后復雜的神經 - 免疫相互作用引起的。由于實驗數據足以描述在時間進程上致病基因的變化(圖 8D),實驗人員還強調了 HMGA2 基因與腸道扭轉不良有關。同樣,早期抑制性運動神經元特異性 NXN 的致病性變異(圖 8Eii) 可導致臍膨出。到該發育進程時,腸道回到了腹部,實驗的結果強調了腸神經細胞和肌肉祖細胞在這個過程中可能是至關重要的。
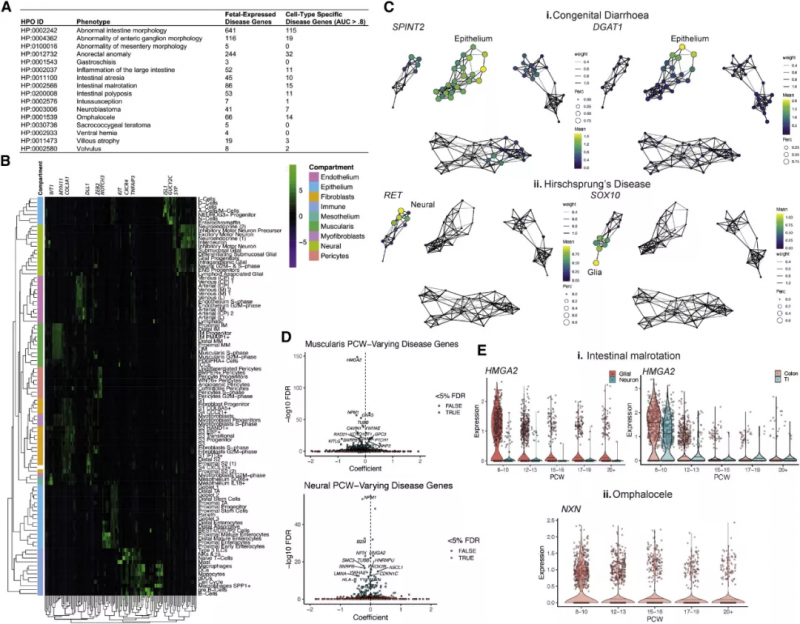
圖 8 子宮內基因表達譜在發育性疾病中的應用
總結
參考文獻:
[1].Fawkner-Corbett D, Antanaviciute A, Parikh K, Jagielowicz M, Gerós AS, Gupta T, Ashley N, Khamis D, Fowler D, Morrissey E, Cunningham C, Johnson PRV, Koohy H, Simmons A. Spatiotemporal analysis of human intestinal development at single-cell resolution. Cell. 2021 Feb 4;184(3):810-826.e23. doi: 10.1016/j.cell.2020.12.016. Epub 2021 Jan 5. PMID: 33406409; PMCID: PMC7864098.
